
Регенераторный ответ гепатоцитов при диффузном токсическом повреждении
УДК 616.36-003.93
З.А. Шафигуллина¹, И.Ф. Гетте¹, И.Г. Данилова¹,²
¹ ФГБУН Институт иммунологии и физиологии УрО РАН, г. Екатеринбург, Российская Федерация;
² ГАУЗ СО Институт медицинский клеточных технологий, г. Екатеринбург, Российская Федерация
Резюме. Изучение регенерации печени при ее токсическом повреждении с целью продления резервов функционирования гепатоцитов является важной задачей. Цель: выявить особенности регенераторной стратегии гепатоцитов при диффузном токсическом повреждении. Материалы и методы. Токсическое повреждение печени у крыс-самцов линии Wistar вызывали путем однократной внутрибрюшинной инъекции масляного раствора тетрахлорметана (CCl₄) в дозе 50 мг/100 г. Животных выводили из эксперимента на 3 и 14 сутки. В рамках проведенного исследования были применены биохимические, иммуногистохимические методы исследования, световая и конфокальная микроскопия, статистический анализ. Результаты. Установлено, что в ответ на действие гепатотропного яда (ССl₄) на 3 сутки наряду с деструктивными процессами (структурные нарушения, усиление апоптоза, дисбаланс биохимических показателей) отмечается увеличение числа пролиферирующих и двуядерных гепатоцитов, а также количествa HSP70+ гепатоцитов. К 14 суткам после введения токсиканта восстановительные процессы реализуются в основном за счет внутриклеточной регенерации, о чем свидетельствует увеличение количества двуядерных гепатоцитов. Нормализуются биохимические показатели, однако при пониженной экспрессии HSP60 в гепатоцитах усиливается апоптотическая гибель клеток. Заключение. Особенности регенераторной стратегии гепатоцитов на ранних сроках после воздействия гепатотропного яда проявляются в увеличении митотической активности паренхиматозных клеток и одновременной активизацией внутриклеточных восстановительных процессов, сопровождающихся увеличением двуядерных клеток и повышением метаболической активности сохранившихся гепатоцитов. Преобладающим типом регенерации в отдаленные сроки, на 14 сутки после внутрибрюшинного введения CCl₄, является внутриклеточная регенерация, благодаря которой стабилизируется функциональная активность гепатоцитов и биохимические показатели крови. Регенераторный ответ гепатоцитов также выражается в изменении продукции белков теплового шока 60 и 70 кДа. Увеличение количества гепатоцитов с содержанием гранул HSP70 на ранних сроках воздействия токсиканта может служить фактором, сдерживающим апоптоз гепатоцитов, в то время как уменьшение HSP60⁺ гепатоцитов напротив служит апоптотическим стимулом.
Ключевые слова: регенерация, гепатоцит, печень, апоптоз, белки теплового шока, токсическое повреждение
Конфликт интересов отсутствует.
Контактная информация автора, ответственного за переписку:
Шафигуллина Злата Александровна
zlata_pyankova@mail.ru
Дата поступления 14.12.2020г.
Образец цитирования:
Шафигуллина З.А., Гетте И.Ф., Данилова И.Г. Регенераторный ответ гепатоцитов при диффузном токсическом повреждении. http://vestnikural.ru/article/1153 [Электронный ресурс] Вестник уральской медицинской академической науки. 2020, Том 17, №4, с. 313–322, DOI: 10.22138/2500-0918-2020-17-4-313-322
Введение
Токсический экспериментальный гепатит, развивающийся при введении CCl₄, является одной из классических моделей для исследования механизмов репаративной регенерации печени [1]. Повреждение печени при действии гепатотропных ядов характеризуется рядом особенностей, что в большей мере определяется структурно-функциональной неоднородностью органа. Гепатоциты перевенозной зоны по сравнению с другими клетками печеночной дольки обладают высокой активностью изоферментов P-450, в связи с чем преимущественно они ответственны за биотрансформацию ядов, вследствие которой образуется большое количество реакционно-активных метаболитов [2]. Ферментные системы, обеспечивающие конъюгацию и антиоксидантную защиту, в этих клетках менее развиты. Поэтому повреждение тетрахлорметаном имеет центролобулярную локализацию и затрагивает гепатоциты перевенозной зоны [3].
Сохранность неповрежденных гепатоцитов в условиях повышенной ксенобиотической нагрузки во многом зависит от целого ряда факторов, к числу которых относится: соотношение процессов апоптоза и пролиферации, функциональная активность, проявляющаяся в секреции ферментов и цитопротекторных белков. В ряде работ показано, что при воздействии неблагоприятных факторов, включая токсичные вещества, усиливается продукция антиапоптотических белков теплового шока [4]. Наибольший интерес представляют белки теплового шока с молекулярной массой 60 и 70 кДа, которые могут действовать как шапероны, возвращая другим белкам нативную конформацию. В частности, HSP 70 начинает процесс сворачивания белков (фолдинг), избыточная продукция HSP 70 защищает митохондрии от воздействия реактивных форм кислорода и является посредником в уменьшении выработки проапоптического фактора — каспазы-3, а комплекс HSP 60 завершает фолдинг [5, 6].
Таким образом, наличие знаний о степени деструктивных изменений ткани печени, преобладающем типе регенерации и количественном изменении HSP 60+ и HSP 70⁺-гепатоцитов на ранних и поздних стадиях токсического повреждения печени могут быть полезны при разработке методов коррекции нарушенных функций гепатоцитов.
Цель исследования: выявить особенности регенераторного ответа гепатоцитов в ранние сроки диффузного токсического повреждения печени.
Материалы и методы
Эксперимент был выполнен на 40 половозрелых (3 месяца) крысах-самцах линии Wistar массой 180±10 г в соответствии с принципами международных этических комитетов (Директива Совета ЕС 2010/63/EU), одобрен комиссией по биоэтике Института естественных наук и математики Уральского федерального университета имени первого Президента России Б. Н. Ельцина (протокол №2 от 21.10.2020 г.) и этическим комитетом Института иммунологии и физиологии УрО РАН (протокол №01/19 от 18.12.2019 г.). Животные, используемые в исследовании, содержались в одинаковых условиях по 5 крыс в клетке на обычном рационе вивария со свободным доступом к пище и воде и температурным режимом 20±2°С.
Для создания модели токсического повреждения печени использовали 50% масляный раствор тетрахлорметана (ССl₄, четырёххлористый углерод, ГОСТ 20288-74, АО «ЭКОС-1»), который вводили животным экспериментальных групп однократно внутрибрюшинно в дозе 50 мг/100 г массы тела, что соответствует 0,3 мл/кг [7]. Продолжительность действия гепатотоксиканта составляла 3 и 14 суток. К группам с CCl₄-воздействием была сформирована контрольная группа животных, которой вводили аналогичную дозу масляного раствора. Интактную группу составляли здоровые животные.
Животных выводили из эксперимента после 12-ти часового голодания передозировкой золетила (15 мг/кг), предварительно взяв кровь из хвостовой вены. После проведения срединной лапаротомии у животных извлекали печень. Фрагменты печени фиксировали в 10%-ном забуференном водном растворе формалина в течение 24 часов. После 8-и часовой промывки фрагменты органов подвергали стандартной гистологической проводке в автоматизированном тканевом процессоре Leica TP 1020 (Leica Microsystems, Германия) с последующей заливкой материала в парафин в системе Leica EG 1160 (Leica Microsystems, Германия). Срезы толщиной 3-4 мкм изготавливали на микротоме Leica SM 2000R (Leica Microsystems, Германия).
Биохимическое исследование плазмы крови крыс включало определение активности аланинаминотрансферазы (АЛТ) (К.Ф.2.6.1.2), аспартатаминотрансферазы (АСТ) (К.Ф.2.6.1.1), щелочной фосфатазы (К.Ф.3.1.3.1.), содержание общего билирубина, общего белка, альбумина, мочевины и креатинина с использованием стандартных наборов реактивов фирмы Витал Диагностикс (Санкт-Петербург, Россия) на приборе DU 800 Beckman Coulter (USA).
Для гистологического и морфометрического исследования ткань печени окрашивали гематоксилином и эозином. Анализ гистологических препаратов проводили с использованием светового микроскопа Leica DM2500 и программного обеспечения для захвата и анализа изображений Leica Application Suite. Определяли митотический индекс (‰), количество двуядерных гепатоцитов в 1 мм² среза. Подсчет вышеперечисленных параметров производился при увеличении микроскопа ×400 в 40 полях зрения.
Иммуногистохимическое окрашивание применялось для идентификации пролиферирующих гепатоцитов (по экспрессии Ki-67). В качестве первичных антител были использованы Purified Mouse Anti-Human Ki67 клон B 56. BD Pharmingentm, USA, авторичных — Biotin Goat Anti-Mouse Ig (Multiple Adsorption), поликлональное. BD Biosciences, USA. Антигенреактивныеклеткивыявлялиприпомощитест-системы NovolinkTM Polymer Detection System (Novocastra Lab., Ltd), контрастируяиххромогеннымсубстратом (3,3-диаминобензидинвбуферномрастворе). DAB-позитивные клетки идентифицировали по коричневому окрашиванию ядер. Количественную оценку Ki-67⁺ гепатоцитов проводили в 40 полях зрения при увеличении микроскопа ×400 с последующим пересчетом на 1 мм² среза.
Для оценки степени апоптоза применяли метод TUNEL (маркировка концевых участков dUTP терминальной дезоксинуклеотидилтрансферазы) с использованием Click-iT Plus TUNEL Assay for in situ apoptosis detection with Alexa Fluor dyes, сat. no. C10618, Thermo Fisher Scientific Inc. Визуализацию и подсчет TUNEL-положительных гепатоцитов осуществляли с помощью лазерного сканирующего конфокального микроскопа LSM 710 (ZEISS, Germany).
Клетки, экспрессирующие белки теплового шока НSP60 и НSP70 в печени определяли иммуногистохимическим (ИГХ) непрямым пероксидазным методом. В ходе ИГХ-окрашивания применяли первичные моноклональные антитела Mouse anti shock protein 60 (hsp60) clone LK1, Merck Millipore, USA, HSP 70 (ЗАЗ): sc-32239, clone 3A3 Santa Cruz Biotechnology, Inc., USA и вторичные антитела Biotin Goat Anti-Mouse Ig (Multiple Adsorption), поликлональное, BD Biosciences, USA. Для исключения неспецифического окрашивания проводили постановку негативного и позитивного изотипического контроля.
Статистическая обработка результатов исследования проводилась с помощью программного обеспечения OriginPro 2018 software (OriginLab Corporation, USA) при помощи непараметрического критерия Краскела-Уоллеса для множественных сравнений (непараметрический метод сравнения трех или более групп). Данный статистический подход был выбран в связи с отсутствием нормального распределения в исследуемых выборках. При проверке статистических гипотез использовался уровень значимости 5% (p<0,05).
Результаты и обсуждение
В ходе проведенного гистологического анализа установлено, что строение печени интактных животных соответствует гистологической норме.
На 3 сутки после введения CCl₄ в ткани печени были выявлены признаки, характерные для острого токсического гепатита (очаговые некрозы гепатоцитов с перифокальной лимфо-лейкоцитарной инфильтрацией), также обнаружены выраженная диффузная вакуольная дистрофия гепатоцитов, анизоцитоз, анизонуклеоз (Рисунок 1) и нарушения микроциркуляторного русла.
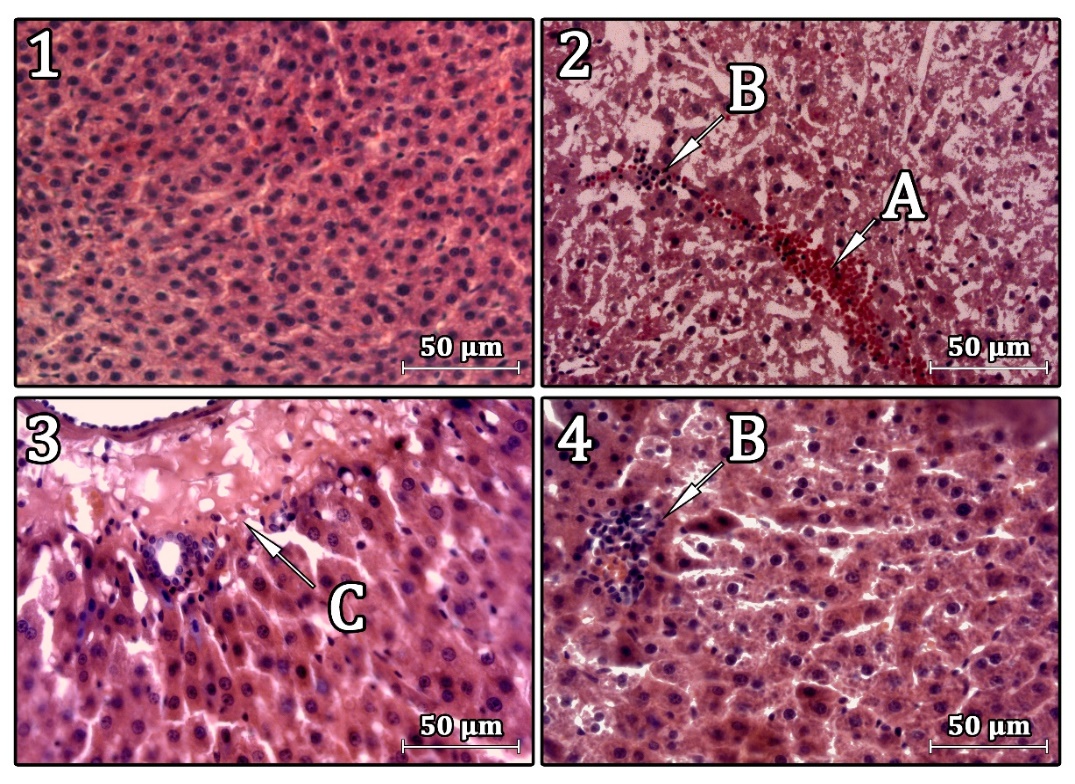
Рисунок 1. Структура печени экспериментальных животных. Окраска гематоксилином и эозином, ув. ×200.
Примечание: 1 – интактная группа; 2 – CCl₄ 3 сутки, 3 и 4 – CCl₄ 14 сутки; А – полнокровие, B – очаговая лимфо-лейкоцитарная инфильтрация, С – некроз гепатоцитов.
На 14 сутки после токсического воздействия CCl₄ балочное строение печени не нарушено. Отмечается очаговое полнокровие вен портальных трактов. Характерна умеренная инфильтрация в области портальных трактов лимфоидными элементами без прорыва пограничной пластинки. Перипортально и в системе междолевых протоков отмечаются некрозы гепатоцитов. Признаки анизоцитоза и анизонуклеоза умеренно выражены, визуально отмечается большое количество двуядерных гепатоцитов. Выявлены признаки диффузной зернистой дистрофии гепатоцитов как следствие нарушения белкового обмена. В синусоидальных пространствах определяются сегментоядерные лейкоциты. Мелкие очаговые инфильтраты в паренхиме органа представлены лимфоидными элементами (Рисунок 1).
Детекция апоптоза показала наличие TUNEL-позитивных клеток во всех исследуемых микропрепаратах печени экспериментальных животных. У интактных крыс TUNEL-позитивные гепатоциты составляют 5,39±0,84 кл/мм² (Рисунок 2). Результаты настоящего исследования подтверждают тот факт, что CCl₄-интоксикация приводит к апоптозу клеток паренхимы печени [6, 8]. На 3 сутки после действия гепатотропного яда отмечается тенденция к увеличению числа TUNEL-позитивных гепатоцитов, что свидетельствует об усилении гибели клеток путем апоптоза. В более поздние сроки исследования, на 14 сутки, происходит увеличение числа апоптотических клеток (Рисунок 2) в ткани печени экспериментальных животных по сравнению с показателями интактной группы.
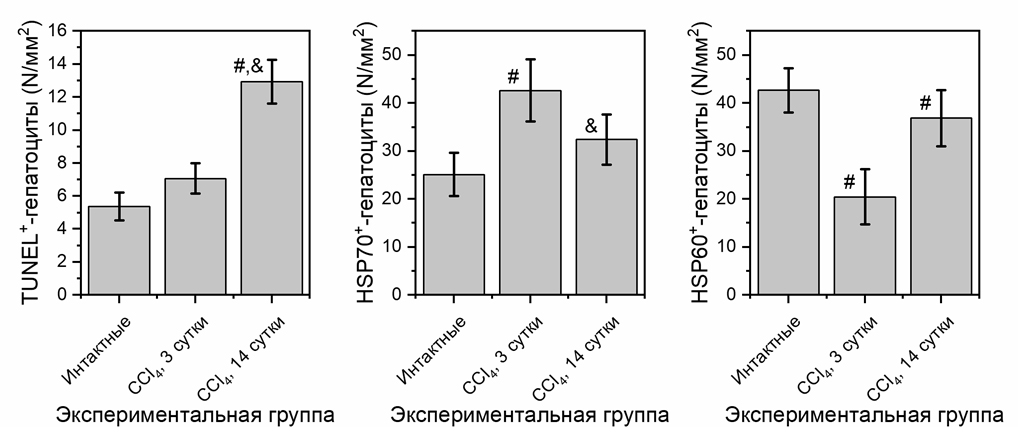
Рисунок 2. Степень апоптоза и количественные изменения гепатоцитов с содержанием гранул белков теплового шока (HSP70 и HSP60)
Примечание: результаты статистически значимы по сравнению с интактной группой (#, р<0,05) и с группой CCl₄ 3 сутки (&, р<0,05).
Структурные изменения печени при токсическом повреждении подтверждаются дисбалансом биохимических показателей (Таблица 1). Введение CCl₄ оказывает существенное влияние на активность ферментов, местом локализации которых в норме являются гепатоциты. На 3 сутки токсического воздействия обнаружено достоверное повышение активности сывороточного АЛТ и АСТ (Таблица 1). Увеличение активности АЛТ, содержащегося в гепатоцитах в большем количестве по сравнению с АСТ, свидетельствует о явлении цитолиза гепатоцитов. Дополнительным биохимическим критерием, подтверждающим повреждение гепатоцитов, является достоверное снижение коэффициента де Ритиса ниже единицы на 3 сутки эксперимента (Таблица 1).
Таблица 1
Биохимические показатели экспериментальных животных в динамике диффузного токсического повреждения
Показатель/Экспериментальная группа | Интактная | CCl₄, 3 сутки | CCl₄, 14 сутки |
АСТ, мкмоль/мин∙л | 16,46±0,96 | 20,52±0,32 # | 16,12±0,75 & |
АЛТ, мкмоль/мин∙л | 12,93±0,88 | 21,04±0,68 # | 12,40±0,57 & |
АСТ/АЛТ, усл. ед. | 1,29±0,06 | 0,98±0,03 # | 1,31±0,06 & |
Щелочная фосфатаза, мкмоль/мин∙л | 69,7±3,9 | 128,2±2,9 # | 76,0±8,1 & |
Мочевина, ммоль/л | 5,09±0,30 | 7,98±0,38 # | 5,42±0,52 & |
Креатинин, мкмоль/л | 64,12±2,45 | 78,95±6,58 | 67,46±4,13 |
Общий белок, г/л | 70,41±2,01 | 69,44±1,76 | 68,70±2,33 |
Альбумины, г/л | 38,59±1,12 | 32,92±0,85 # | 34,26±1,53 & |
Глобулины, г/л | 31,83±1,98 | 36,52±2,16 | 34,44±2,16 |
Альбумины/Глобулины, усл. ед. | 1,25±0,08 | 0,91±0,06 # | 1,02±0,09 & |
Общий билирубин, мкмоль/л | 2,71±0,25 | 17,05±2,27 # | 2,93±0,15 & |
Примечание: результаты статистически значимы по сравнению с интактной группой (#, р<0,05) и с группой CCl₄ 3 сутки (&, р<0,05).
В эти же сроки наблюдается повышение активности щелочной фосфатазы. Этот фермент локализован на мембране гепатоцитов, обращенной в сторону желчного капилляра, и в норме должен экскретироваться в желчь [9]. Увеличение активности щелочной фосфатазы в плазме крови свидетельствует о развитии холестатического синдрома, при котором компоненты желчи задерживаются в печени и при нарушении целостности мембран гепатоцитов поступают в кровь. Развитие холестатического синдрома подтверждается увеличением количества билирубина также на 3 сутки после введения CCl₄ (Таблица 1).
При сравнении содержания общего количества белка в группах крыс на 3 и 14 сутки после введения токсиканта не было обнаружено достоверных отличий по сравнению с показателем интактной группы. В то же время было выявлено снижение содержания альбуминов в плазме крови на 3 сутки эксперимента (Таблица 1). Уменьшение количества альбуминов в плазме крови указывает на недостаточность белоксинтетической функции печени, что подтверждается снижением альбумино-глобулинового индекса на ранний срок после введения CCl₄ (Таблица 1).
Известно, что действие тетрахлорметана сопровождается поражением почек [10]. Следует отметить увеличение уровня мочевины на 3 сутки после введения CCl₄ (Таблица 1) при неизменном содержании сывороточного креатинина, что можно расценивать как проявление в небольшой степени нарушения фильтрации в почечных клубочках.
Данные биохимического анализа подтверждают, что при действии CCl₄ в дозе 50 мг/100 г массы тела у экспериментальных животных на 3 сутки развивается токсический гепатит, сопровождающийся развитием цитолиза, внутреннего холестаза, умеренным снижением синтетической функции печени и процесса фильтрации в почках. Через 14 суток с момента введения гепатотропного яда биохимические показатели снижаются относительно параметров группы CCl₄ 3 сутки и соответствуют уровню интактных.
При повреждении паренхимы печени CCl₄ активизируются адаптивно-компенсаторные механизмы. После действия гепатотропного яда CCl₄ в печени экспериментальных животных на 3 сутки резко возрастает количество двуядерных гепатоцитов до 51,8±0,3 кл/мм² по сравнению с показателями интактной группы 15,4±1,4 кл/мм² (Рисунок 3). Через две недели после внутрибрюшинного введения CCl₄ количество двуядерных гепатоцитов возросло до 64,27±5,37 кл/мм², что существенно превышает показатели интактных животных и на 24% выше (Рисунок 3), чем в экспериментальной группе на 3 сутки.
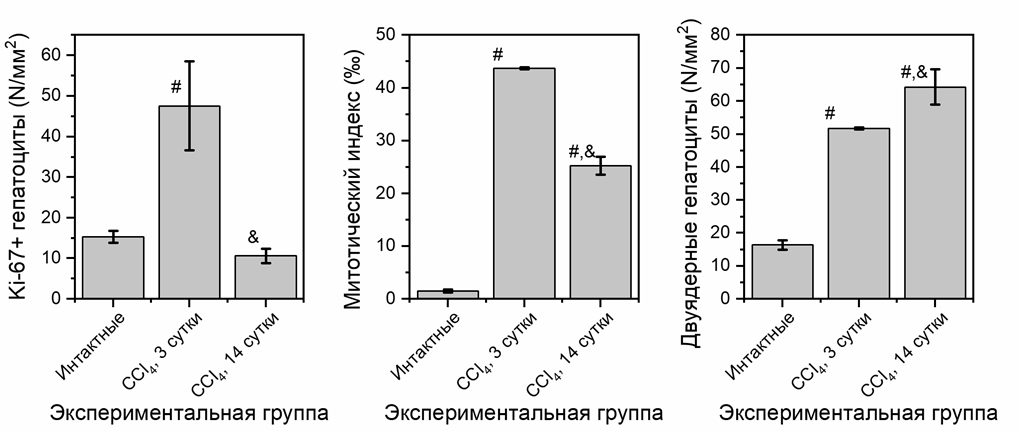
Рисунок 3. Показатели регенерации печени в норме и при диффузном токсическом повреждении
Примечание: результаты статистически значимы по сравнению с интактной группой (#, р<0,05) и с группой CCl₄ 3 сутки (&, р<0,05).
Кроме процессов внутриклеточной регенерации при диффузном токсическом повреждении печени в ранние сроки наблюдается активация клеточной регенерации. На это указывает увеличение митотического индекса и количества пролиферирующих Ki-67⁺-гепатоцитов (Рисунок 3). К 14 суткам митотический индекс снижается относительно группы CCl₄, 3 сутки на 42%, но при этом значительно выше, чем у интактных животных. Количество Ki-67⁺ гепатоцитов на 14 сутки имеет тенденцию к снижению (Рисунок 3), однако достоверных различий не выявлено.
Таким образом, в ранние сроки токсического воздействия CCl₄ ответная реакция паренхимы печени выражается в виде усиления клеточной и внутриклеточной регенерации. Данная стратегия способствует повышению метаболической активности, выражающейся в увеличении количества гепатоцитов с содержанием гранул HSP 70 на 3 сутки эксперимента (Рисунок 2). Именно HSP70 участвуют в начальных этапах восстановления нативной конформации белков и служат инициаторами регенерации. Повышенная продукция HSP 70, вероятно, защищает клетку от воздействия токсиканта и продуктов метаболизма, а также препятствует переходу начальных реакций апоптоза в необратимую стадию [11].
При воздействии гепатопропного яда существенно снижается количество HSP60⁺ гепатоцитов как на 3, так и на 14 сутки эксперимента (Рисунок 3), при этом, как упоминалось ранее, возрастает степень апоптоза.
Вероятно, продукция гепатоцитами белков теплового шока 60 и 70 кДа определяет соотношение процессов пролиферации и клеточной гибели, в связи с чем может рассматриваться в качестве потенциального фактора для коррекции нарушений печени.
Заключение
Особенности регенераторного ответа гепатоцитов на ранних сроках после воздействия гепатотропного яда проявляются в увеличении митотической активности паренхиматозных клеток и одновременной активизацией внутриклеточных восстановительных процессов, сопровождающихся увеличением двуядерных клеток и повышением метаболической активности сохранившихся гепатоцитов. Преобладающим типом регенерации в отдаленные сроки, на 14 сутки после внутрибрюшинного введения CCl₄, является внутриклеточная регенерация, благодаря которой биохимические показатели приходят в норму.
Регенераторный ответ гепатоцитов также выражается в изменении продукции белков теплового шока 60 и 70 кДа. Увеличение количества гепатоцитов с содержанием гранул HSP70 на ранних сроках воздействия токсиканта может служить фактором, сдерживающим апоптоз гепатоцитов, в то время как уменьшение HSP60⁺ гепатоцитов напротив служит апоптотическим стимулом.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 19-315-90012 и бюджетной программы № Гос. регистрации – АААА-А18-118020590107-0 с использованием оборудования ЦКП ИИФ УрО РАН.
ЛИТЕРАТУРА
- Forbes S., Newsome P. Liver regeneration – mechanisms and models to clinical application. Nat Rev Gastroenterol Hepatol. 2016, 13: 473–485. doi: 10.1038/nrgastro.2016.97.
- Gu X., Manautou J. E. Molecular mechanisms underlying chemical liver injury. Expert Rev Mol Med. 2012, 14: e4. doi:10.1017/S1462399411002110.
- Khan A. A., Alzohairy M. A. Hepatoprotective effects of camel milk against CCl₄ induced hepatotoxicity in rats. Asian Journal of Biochemistry. 2011, 6: 171–180. doi: 10.3923/ajb.2011.171.180.
- Mahmood K., Jadoon S., Mahmood Q., Irshad M., Hussain J. Synergistic effects of toxic elements on heat shock proteins. BioMed research international. 2014: 564136. doi: 10.1155/2014/564136.
- Walter S., Buchner J. Molecular chaperones-cellular machines for protein folding. Angew Chem Int Ed Engl. 2002. 41(7): 1098–113.
- Shi J., Aisaki K., Ikawa Y., Wake K. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am J Pathol. 1998. 153: 515–525.
- Руководство по проведению доклинических исследований лекарственных средств. Часть первая. М.: Гриф и К, 2012. 944 с.
- Zahedi K., Barone S. L., Xu J., Steinbergs N., Schuster R., Lentsch A. B. et al. Hepatocyte-specific ablation of spermine/spermidine-N1-acetyltransferase gene reduces the severity of CCl₄-induced acute liver injury. Am J Physiol Gastrointest Liver Physiol. 2012. 303: 546–560.
- ТагановичА. Д. идр. Биологическая химия: 2-е изд., исправленое. Минск: Вышэйная школа, 2016. 671 с.
- Калетина Н. И. ред. Токсикологическая химия. Метаболизм и анализ токсикантов: учебное пособие. М.: ГЭОТАР-Медиа, 2008. 1016 с.
- Kurashova N. A., Madaeva I. M., Kolesnikova L. I. Expression of HSP70 Heat-Shock Proteins under Oxidative Stress. Adv Gerontol. 2020. 10: 20–25. doi: 10.1134/S2079057020010099.
Авторы
Шафигуллина Злата Александровна
ФГБУН Институт иммунологии и физиологии УрО РАН
Младший научный сотрудник лаборатории морфологии и биохимии
Российская Федерация, 620049, г. Екатеринбург, ул. Первомайская, д. 106
zlata_pyankova@mail.ru
Гетте Ирина Федоровна
ФГБУН Институт иммунологии и физиологии УрО РАН
Кандидат биологических наук, старший научный сотрудник лаборатории морфологии и биохимии
Российская Федерация, 620049, г. Екатеринбург, ул. Первомайская, д. 106
i.goette@yandex.ru
Данилова Ирина Георгиевна
ФГБУН Институт иммунологии и физиологии УрО РАН
Доктор биологических наук, доцент, заведующая лабораторией морфологии и биохимии
ГАУЗ СО Институт медицинских клеточных технологий
Главный научный сотрудник
Российская Федерация, 620026, г. Екатеринбург, ул. К. Маркса, д. 22а
ig-danilova@yandex.ru